

01.03.2022 - 04:01
L’ADN és la biomolècula fonamental per al correcte funcionament cel·lular i fins fa poc s’ha associat a l’estructura de doble hèlix descoberta fa més de seixanta anys per Crick, Watson i Franklin. Tanmateix, s’han descrit altres estructures i conformacions diferents que l’ADN pot adoptar com els G-quàdruplex. Aquests G-quàdruplex es formen en regions del genoma riques en guanines amb una estructura tetràmera i controlen processos biològics com l’expressió genètica, la protecció contra l’envelliment o la transmissió de la informació neuronal. En aquest article descrivim les seues característiques químiques i estructurals a més d’introduir les seues principals funcions cel·lulars. Per finalitzar, presentem els G-quàdruplex com a dianes moleculars per a futures teràpies contra el càncer.
Avui dia es coneix que els àcids nucleics són biomolècules fonamentals per al correcte funcionament cel·lular i la regulació dels processos biològics. Aquestes biomolècules poden plegar-se en diverses estructures i conformacions. Tanmateix, l’estructura més coneguda és l’obtinguda per Rosalind Franklin, James Watson i Francis Crick als anys cinquanta, la famosa doble hèlix de l’àcid desoxiribonucleic o ADN. El descobriment de l’estructura de la molècula de la vida va revolucionar la biologia molecular del segle XX perquè servia per a comprendre com s’emmagatzemava, protegia i transmetria la informació genètica dels éssers vius. L’ADN és una estructura polimèrica helicoidal composta per quatre nucleòtids que es diferencien entre si per la base nitrogenada que contenen: adenina (A), guanina (G), timina (T) o citosina (C). Així doncs, l’ADN es pot entendre com un codi de lletres que actua com a guia per a la construcció de l’àcid ribonucleic o ARN, el qual serveix, majoritàriament, per a generar les proteïnes que regulen totes les funcions biològiques i característiques dels éssers vius, com ara el color d’ulls, la forma dels cabells o la grandària de les mans.
A banda de l’estructura en doble hèlix, s’ha descobert que l’ADN pot formar conformacions alternatives que controlen diverses funcions biològiques, com ara l’expressió gènica o la protecció contra l’envelliment. Entre aquestes estructures, l’ADN G-quàdruplex (G4) és la que més atenció ha rebut en els últims deu anys a causa del seu rol com a regulador epigenètic i com a diana terapèutica de malalties com el càncer. En aquest article volem introduir els lectors en aquesta nova i magnífica estructura d’ADN, tant pel que fa a l’estructura com a les seues funcions moleculars i biològiques. A més a més, analitzem les seues aplicacions potencials com a diana terapèutica en diverses malalties i expliquem les perspectives futures d’investigació en aquest camp.
La diversitat estructural de l’ADN
La cursa pel descobriment de l’estructura de l’ADN va culminar amb els experiments de difracció de fibres duts a terme per Rosalind Franklin, que donaren lloc a l’obtenció de la famosa Imatge 51 de raigs X. Tanmateix, James Watson i Francis Crick van publicar la descripció de l’estructura de la doble hèlix en la revista Nature basant-se en aquesta imatge (Watson i Crick, 1953) i anys després, el 1962, ambdós, junt amb Maurice Wilkins, van ser guardonats amb el Premi Nobel de Fisiologia i Medicina (Oregon State University, 2015). Aquest descobriment va esdevenir l’avanç més significatiu i extraordinari de la biologia molecular i una de les troballes científiques més importants del segle XX.
L’estructura de doble hèlix d’ADN es compon de dues cadenes enfrontades entre si de forma antiparal·lela formant una espiral que crea una estructura helicoidal. Com ja s’ha comentat abans, les cadenes d’ADN es formen a partir de quatre unitats distintes, els nucleòtids. Els nucleòtids inclouen un grup fosfat, un sucre i una base nitrogenada. Les bases nitrogenades són l’adenina, guanina, timina i citosina. Pel que fa als grups fosfats, aquests s’orienten cap a l’exterior de l’estructura, mentre que les bases s’allotgen en l’interior, i donen lloc a parells de bases enfrontades (guanina-citosina i adenina-timina) que es troben unides per enllaços d’hidrogen. Cada parell de bases es col·loca damunt d’un altre amb una rotació determinada. Així, es forma l’espiral en l’eix central de l’estructura que molts artistes, escultors i arquitectes han utilitzat per crear les seues obres. A més, cal comentar que cadascuna de les cadenes d’un àcid nucleic té dos extrems a què ens referim com a extrem 5’ i extrem 3’. Així, podem assignar una orientació o direccionalitat de l’extrem 5’ a l’extrem 3’ per a descriure els processos que ocorren en l’ADN.
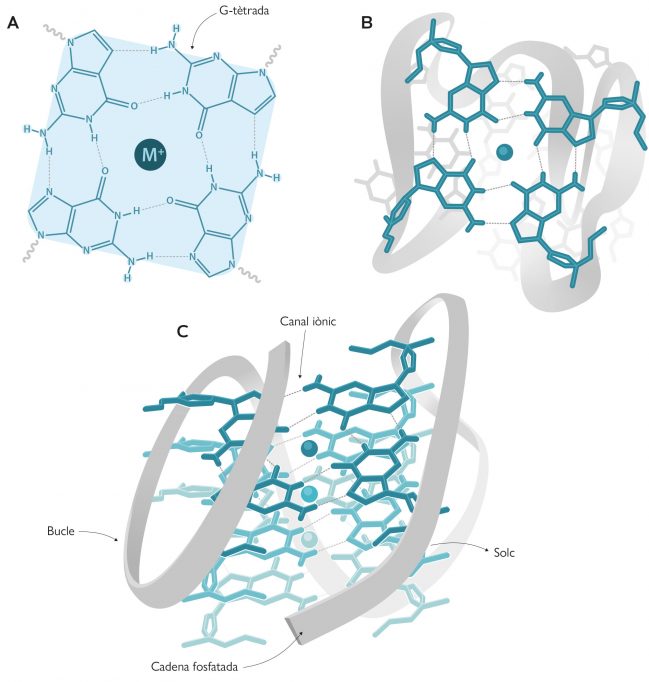
Tanmateix, l’estructura en doble hèlix no és l’única que es coneix en l’actualitat per a l’ADN. Hi ha altres formes no canòniques, com ara els ADN-tríplex (ADN de triple cadena), els i-motif (semblants als G-quàdruplex) o les forquetes de replicació, que fan de l’ADN una molècula amb una gran varietat conformacional. De fet, uns deu anys més tard del desxiframent de l’estructura de l’ADN, Martin Gellert, David R. Davies i col·laboradors publicaren les primeres estructures compostes per gels de guanines que formaven G-tètrades (Gellert et al., 1962), les unitats bàsiques del G-quàdruplex. Aquestes G-tètrades, anomenades també G-quartets, contenen quatre guanines unides per enllaços d’hidrogen, que adopten una disposició plana (Figura 1a-b). Però no va ser fins als anys vuitanta quan Elizabeth H. Blackburn i el seu equip van descriure la primera estructura G-quàdruplex formada per seqüències d’ADN en els telòmers (Henderson et al., 1987). Aquest descobriment va despertar un gran interès en la comunitat científica, no sols per l’estructura en si, sinó per les seues implicacions biològiques. Açò va fer que ràpidament augmentés el nombre d’estudis per a esbrinar quines regions del genoma humà podien formar aquestes estructures. Mitjançant tècniques bioinformàtiques i l’anàlisi de seqüenciació genòmica es van localitzar 700.000 regions en el genoma humà que podrien formar G-quàdruplexs (Chambers et al., 2015). Però el més extraordinari de tot és que no es localitzaven a l’atzar, com era d’esperar d’una regió sense funcionalitat, sinó que es detectaven principalment en els telòmers i en els promotors d’oncogens. Aquest fet indica que els G-quàdruplex tenen un paper primordial en els éssers vius. Com va dir Aaron Klug, premi Nobel de Química per desenvolupar mètodes cristal·logràfics per a desxifrar els complexos proteics amb àcids nucleics, «si els G-quàdruplexs es formen tan fàcilment in vitro, la natura deu haver trobat una manera d’utilitzar-los in vivo».
L’estructura del G-quàdruplex
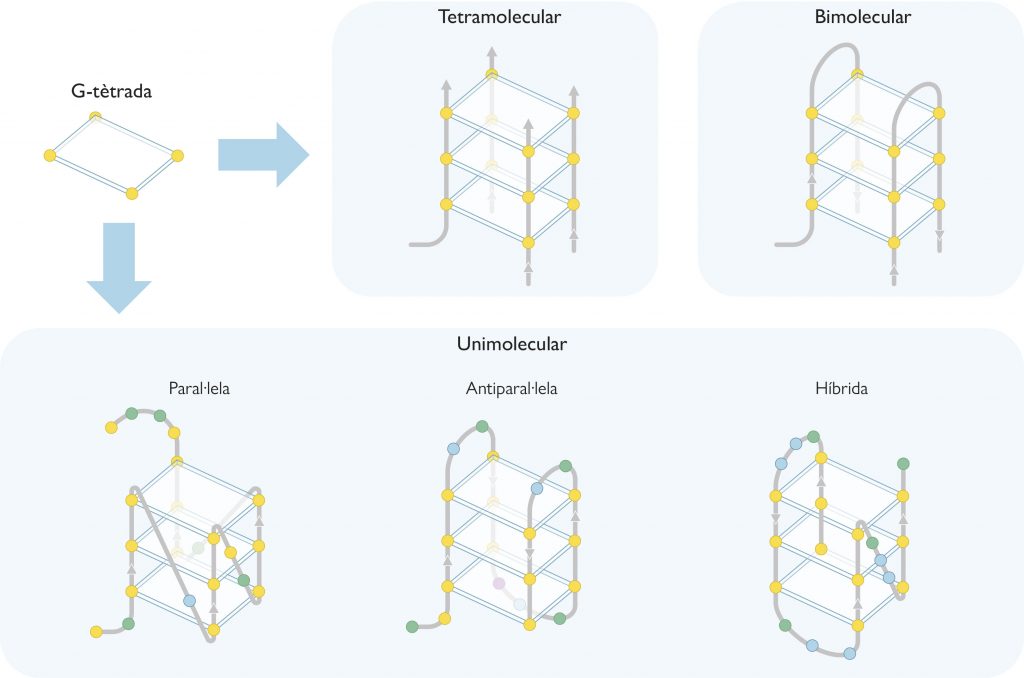
Com ja hem apuntat abans, els G-quàdruplexs estan formats per anells de quatre guanines unides per enllaços d’hidrogen que adopten una disposició plana coneguda com a G-tètrada. Cada G-quàdruplex conté dues o més G-tètrades que s’apilen de forma similar a les plantes d’un edifici a l’interior del qual es crea un mena d’ull d’escala (un canal iònic) que conté cations, normalment de sodi o potassi (Figura 1). Depenent de la grandària del catió, aquest se situa en la mateixa planta de l’edifici (és a dir, al mateix pla que cada G-tètrada) i forma quatre enllaços amb les guanines, com ocorre en el cas del sodi; el potassi, en canvi, és més gran i es col·loca entre dues plantes de l’edifici (és a dir, entre dues G-tètrades) i forma vuit enllaços (Neidle i Balasubramanian, 2006).
Els G-quàdruplexs poden presentar multitud de conformacions regides per diversos paràmetres. Així doncs, poden estar formats per una (quàdruplex unimolecular), dos (bimolecular) o quatre (tetramolecular) cadenes d’ADN riques en guanines (Figura 2). Segons l’orientació relativa d’aquestes cadenes també hi ha diverses conformacions, com ara la paral·lela, l’antiparal·lela i l’híbrida (Figura 2). En la conformació paral·lela totes les cadenes tenen la mateixa orientació, mentre que en l’antiparal·lela, dues presenten una orientació oposada a les altres. Pel que fa a la conformació híbrida, tres cadenes s’orienten en un sentit i la restant, en l’oposat. D’altra banda, aquestes cadenes d’ADN formen bucles i solcs en els G-quàdruplexs (Figura 1c), que depenen de la composició de nucleòtids de les cadenes i d’altres factors externs, com poden ser la força iònica o la temperatura.
El G-quàdruplex en els telòmers
Els telòmers es troben en els extrems dels cromosomes, els quals estan compostos per regions d’ADN no codificant de seqüències altament repetitives. Tenen una funció essencial per a mantenir la integritat dels cromosomes en processos com la recombinació genètica, la fusió de cromosomes i la degradació per nucleases. En definitiva, els telòmers conserven la informació gènica durant les divisions cel·lulars i, per tant, preserven la integritat i funcionalitat dels gens en el temps. Concretament, l’ADN telomèric està compost per una repetició de nucleòtids rica en guanines, que normalment és TTAGGG per a les cèl·lules eucariotes, i que presenta una llargària d’entre 9.000 a 15.000 nucleòtids. Tot i que l’ADN telomèric presenta una conformació de doble hèlix, en el seu extrem 3’ sempre hi ha un fragment d’ADN monocatenari d’entre 50 i 300 bases de llargària, que sobra respecte a la seua cadena complementària (Figura 3).
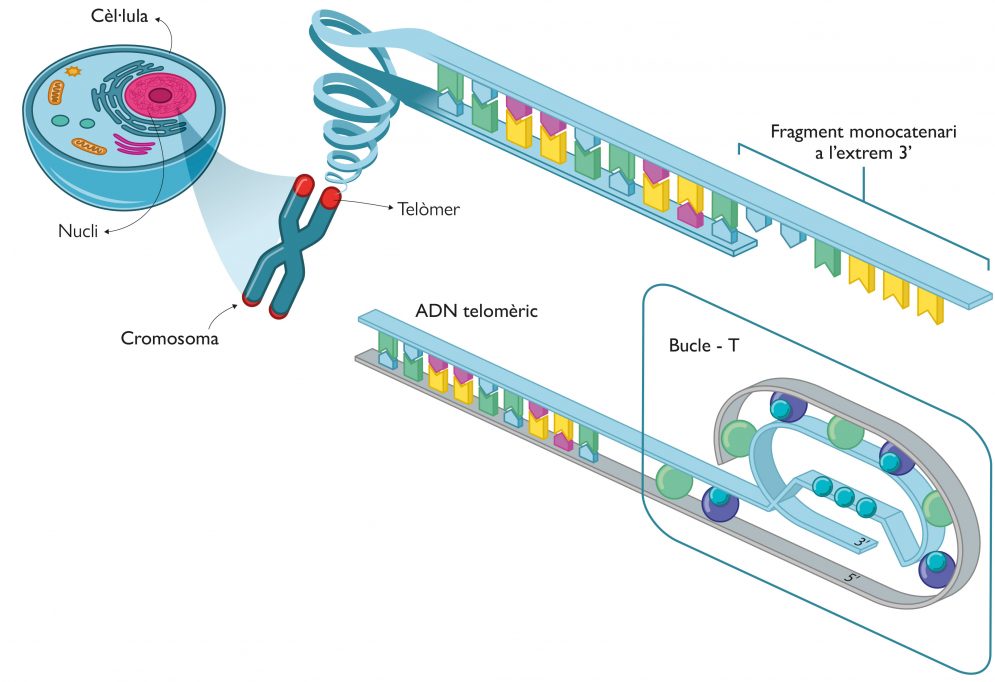
L’ADN monocatenari als telòmers forma una estructura en forma de bucle (coneguda com a bucle-T), en què l’ADN es cargola sobre si mateix i a la qual s’uneixen una sèrie de proteïnes. Aquesta estructura proteica s’anomena supercomplex shelterina o telosoma i protegeix els telòmers de la degradació pròpia de l’ADN que està desbridat en els extrems perquè es troba en forma monocatenària sense emparellar-se amb la brida complementària (Figura 3). Malgrat l’important paper que els telòmers tenen en la protecció dels cromosomes, aquests s’acurten en cada divisió cel·lular a causa del conegut «problema de la replicació», que resulta en la mort cel·lular. Segons el mecanisme de replicació típic de l’ADN, l’enzim polimerasa requereix un petit fragment d’ARN, anomenat encebador, que actua com a iniciador per a afegir nucleòtids a la cadena nova d’ADN que es forma. Com que l’addició sempre és en direcció 5’ a 3’, una de les dues cadenes se sintetitza de forma contínua, mentre que l’altra ha de construir-se de manera discontínua amb fragments curts d’ADN anomenats fragments d’Okazaki, cadascun dels quals té el seu propi encebador que serveix com a motlle per a iniciar l’addició de nucleòtids en la direcció 5’ a 3’. En finalitzar, els fragments d’ARN són eliminats i substituïts per ADN, amb l’excepció de l’últim encebador, el de l’extrem 5’. Aquest és precisament «el problema» i l’origen del fragment monocatenari 3’ dels telòmers. Consegüentment, en cada cicle cel·lular es produeix una pèrdua neta de nucleòtids d’entre 100 a 200 bases. Després d’un determinat nombre de cicles, s’assoleix el conegut com a límit Hayflick, on la longitud dels telòmers és tan curta que ni es pot formar el bucle-T ni tampoc es pot produir un altre cicle. En aquest punt, la cèl·lula té associada una gran inestabilitat genòmica i entra en l’etapa de senescència, en la qual s’atura la divisió i, en última instància, es produeix la mort cel·lular.
Llig ací l’article complet publicat a la Revista Mètode
